http://www.americanarachnology.org/JoA_tocs/JOA_contents_v39n3.html
Shaw, E. M., S. P. Bennett & C. P. Wheater. 2011. Distribution of Brachypelma vagans (Theraphosidae) burrows and their characteristics in Belize over two years. Journal of Arachnology 39 (3): 515-518. doi: http://dx.doi.org/10.1636/P10-39.1
Abstract. To help to address the paucity of ecological knowledge available for Brachypelma vagans Ausserer 1875, a CITES protected species, we monitored a population in Western Belize for two years to provide data for distribution and dispersal. Despite previous over-collection for the pet trade, the species is locally highly abundant in some areas of Belize. We recorded the distribution, burrow and spider characteristics of B. vagans in 2007 and 2008 at Las Cuevas Research Station, Belize. Population dynamics were compared between years, as was individual location. Over 100 burrows were located in both years; however, previous assumptions that individuals do not move burrows regularly appear negated, since only 12 burrow locations matched between years, suggesting high intra-habitat dispersal. Despite this apparent high level of movement burrows were significantly clumped, to a similar degree, in both years. This movement could be due to disturbances throughout the year, including flooding during the rainy season. Burrow size correlated with individual body size, except in a few juveniles that appear to have opportunistically claimed an empty burrow, accounting for some small animals found in large burrows.
Guadanucci, J. P. L. 2011. The genus Plesiophrictus Pocock and revalidation of Heterophrictus Pocock (Araneae: Theraphosidae). Journal of Arachnology 39 (3): 523-527. doi: http://dx.doi.org/10.1636/A10-53.1
Abstract. The genus Plesiophrictus is diagnosed and redescribed based on type material and additional specimens. The type species P. millardi (Pocock 1899) is redescribed. The genus Heterophrictus is revalidated, with H. milleti as type species. Heterophrictus differs from Plesiophrictus by the absence of serrula on maxillae and by having a rastellum on the chelicerae and stiff, spike-shaped setae on the prolateral coxae I. The significance of characters used in the taxonomy of both genera is discussed.
Fukushima, C. S., F. Pérez-Miles & R. Bertani. 2011. On the Avicularia (Araneae: Theraphosidae: Aviculariinae) species from Uruguay. Journal of Arachnology 39 (3) :528-532. doi: http://dx.doi.org/10.1636/A10-12.1
Abstract. The taxonomic status of four species of Avicularia Lamarck 1818 described from Uruguay: Avicularia anthracina (C.L. Koch 1842), Avicularia alticeps (Keyserling 1878), Avicularia parva (Keyserling 1878) and Avicularia tigrina (Pocock 1903) is discussed. The holotypes and/or original descriptions of these species were examined, and two taxonomic synonymies are needed, which are presented herein. Avicularia anthracina is transferred to Grammostola, resulting in Grammostola anthracina (C.L. Koch 1842) new combination and is considered a senior synonym of Grammostola mollicoma Ausserer 1875 new synonymy. Likewise, Avicularia parva is transferred to Catumiri Guadanucci 2004, where it is placed in the synonymy of Catumiri uruguayense Guadanucci 2004 new synonymy. Avicularia tigrina and Avicularia alticeps, originally described in the genera Ischnocolus Ausserer 1875 and Pterinopelma Pocock 1901, respectively, are herein considered nomina dubia since their types are presumed lost.

Monday, December 26, 2011
Wednesday, December 21, 2011
Boldog karácsonyt
Sok szeretettel kívánunk az Apropók blog nevében Boldog Karácsonyt minden kedves olvasónak.
Saturday, December 10, 2011
Hamupipőke
Aki ott volt az idei Pókásztalálkozón, és még emlékszik az előadásomra, az nagyjából sejtheti, hogy miről lesz szó. Hát azóta vannak új fejlemények. Addig is amíg nem írhatok róla (majd ha elfogadták a cikket), akkor majd fogok. :D



Friday, November 04, 2011
Régi, de nem elfeledett
Több, mint 100 év után találtak rá újra egy madárpókfajra. A Biologia Centrali Americana-ban F. O. P.-Cambridge által 1897-ben, Guatemalából leírt Citharacanthus longipes fajból eddig csak az eredeti típuspéldányok voltak ismertek. Az idén nyáron a Cusuco Nemzeti Parkban (Honduras) végzett Operation Wallacea felmérés egyik gyümölcse két, egymás mellett élő madárpókfaj felfedezése volt. A múzeumi példányokkal való összevetés után az egyik faj az elveszettnek hitt C. longipes-nek bizonyult, a másik pedig a (relatíve) közelmúltban leírt C. livingstoni Schmidt and Weinmann, 1996. Habár a második faj azonosítása még nem teljesen biztos, mindkét faj jelenléte Hondurasban új adat/eredmény.
Sunday, October 23, 2011
Tuesday, October 11, 2011
Wednesday, September 07, 2011
Milyen Tina? Bonnetina!
Estrada-Alvarez és Locht a Sociedad Entomológica Aragonesa (S.E.A.) bulletinjében megjelent cikkében egy tudományra új Bonnetina faj leírását olvashatjuk. A Bonnetina aviae-nek nevezett faj négyre növeli a tudomány számára ismert Bonnetina fajok számát. A többi három fajtól az ivarhólyag, spermatartók, a csáprágók kitinfogainak száma, valamint a lábak hosszúságainak aránya alapján különítik el. Az új fajról képet jelenleg nem tudok itt közzétenni, aki viszont nem ismeri a Bonnetina génuszt, Rick West honlapján nézelődhet (és észreveheti, hogy több faj vár még leírásra).
Estrada-Alvarez, J. C. & A. Locht. 2011. Descripción de Bonnetina aviae sp.n. de Méxicó (Araneae: Theraphosidae: Theraphosinae). Boletín de la Sociedad Entomológica Aragonesa (S.E.A.) 48: 151‒155.
Tuesday, September 06, 2011
Stuart Longhorn-ék Hondurasi pókvadászata
Stuart Longhorn nyáron Hondurasban járt, pókszabásúakat gyűjtöttek egy nemzeti park felméréséhez. A madárpókos képek Stuart flickr albumában megtekinthetők.
Sunday, August 28, 2011
A nyáron megjelent madárpókos cikkek, összefoglalókkal
Ferretti, N. E. & F. Pérez-Miles. 2011. Intraspecific non-sexual interactions of Grammostola schulzei (Araneae: Theraphosidae) under laboratory conditions. Revista de Biología Tropical (International Journal of Tropical Biology and Conservation) 59 (3): 1173–1182.
Abstract. Intraspecific interactions of araneomorph spiders have received considerable attention, but there are few detailed studies on intraspecific interactions of mygalomorph spiders. Moreover, a thorough understanding of theraphosid biology and ecology is necessary from a conservation standpoint because natural populations may be threatened by habitat disturbances and captures for pet commerce. We described the behavior of conspecific individuals of Grammostola schulzei during non-sexual interactions, under laboratory conditions. Pairs of individuals involving adult males, adult females and juveniles were confronted and observed in resident and intruder conditions, totalizing 115 trials. When confronted two adult females, they retreated or grappled, and performed gaping display with bite attempts, usually resulted in severe injury of the intruder spiders. When confronted females with large juveniles, we frequently observed cannibalism on juveniles. Juveniles exposed to females or to other juveniles retreated or made leg tapping with forelegs and palpal drumming, which are common displays of courting adult males. Adult males courted and clasped some juveniles, but juveniles avoided or reject clasping. The behaviors observed during intraspecific interactions could play an important role determining spatial distribution and could lead to behavioral adaptations of territoriality. Rev. Biol. Trop. 59 (3): 1173-1182. Epub 2011 September 01.
Megjegyzés: a Grammostola schulzei (Schmidt, 1994) fajt a közelmúltban szinonimizálták a G. vachoni Schiapelli & Gerschman, 1961 fajjal [Ferretti et al. (2011)].
Dor, A., S. Calmé & Y. Hénaut. 2011. Predatory interactions between Centruroides scorpions and the tarantula Brachypelma vagans. Journal of Arachnology 39 (1) :201–204. DOI: 10.1636/St08-84.1
Abstract. In the Yucatan Peninsula, the tarantula Brachypelma vagans Ausserer 1875 is commonly associated with human settlements, as are the scorpions Centruroides gracilis Latreille 1804 and C. ochraceus Pocock 1898. Nonetheless, scorpions are virtually absent from villages showing a high density of tarantulas. Predatory interactions between these predators could explain the lack of local overlap. To test this hypothesis, we observed the behavioral interactions between B. vagans and C. gracilis or C. ochraceus in experimentally controlled conditions, and we compared these interactions to interactions between the tarantula and two prey species: cricket and cockroach. For observations, a pre-adult tarantula was placed in an experimental arena in which we introduced either a scorpion or an insect. In all, 115 trials were performed. We recorded time elapsed and behavioral responses: avoidance, attack, escape, capture, and attack success. Tarantulas preyed on all prey with the same attack success (63.8% ± 0.8%), but they attacked and captured cockroaches quicker and more often than the other prey (87% vs. 50%, and 57% vs. 30%, respectively). Scorpions attacked tarantulas in 25.5% of occasions, but they were never successful, and were killed in 9% of occasions. We conclude that tarantulas are potential predators of scorpions. Moreover, in villages where tarantulas are abundant they might prevent the presence of scorpions. Thus the presence of this non-aggressive tarantula may be beneficial from the human perspective.
Canal, M., D. Figueroa, C. Alfaro, T. Kawamoto, H. Torres-Contreras, P. Sabat & C. Veloso. 2011. Effects of diet and water supply on energy intake and water loss in a mygalomorph spider in a fluctuating environment of the central Andes. Journal of Insect Physiology, Article in Press, Uncorrected Proof. DOI: 10.1016/j.jinsphys.2011.07.016
Abstract. The metabolic and water evaporation strategies in spiders may be part of a set of physiological adaptations to tolerate low or unpredictable food availability, buffering spiders against environmental fluctuations such as those of the high mountains of the central Andes.
The aim of this study is to analyze experimentally the variations in metabolic rate and the rate of evaporative water with food and/or water restriction in a high mountain mygalomorph spider population (Paraphysa sp.).
We found that the low metabolism of this spider was not affected by water restriction, but its metabolism was depressed after 3 weeks of food deprivation. The spider did not show seasonal metabolic changes but it presented seasonal changes in the rate of evaporative water loss at high temperatures.
Females with egg sacs reduced their metabolic rate and evaporative water at high temperatures.
These findings constitute a set of possible adaptations to a highly fluctuating Mediterranean environment, which is completely covered with snow for many months and then progresses rapidly to a very dry climate with high temperatures.
Wang, Z. Y., J. T. Wang, A. H. Ji, H. K. Li & Z. D. Dai. 2011. Movement behavior of a spider on a horizontal surface. Chinese Science Bulletin 56: 2748–2757. DOI: 10.1007/s11434-011-4584-y
Abstract. Studying the locomotive behavior of animals has the potential to inspire the design of the mechanism and gait patterns of robots (“bio-inspired robots”). The kinematics characteristics of a spider (Ornithoctonus [sic] huwena), including movement of the legs, movement of the center of mass (COM) and joint-rotation angle, were obtained from the observation of locomotion behaviors recorded by a three-dimensional locomotion observation system. Our results showed that one set of the stance phase consists of four legs, which were leg-1 and leg-3 on one side and leg-2 and leg-4 on the other side. Additionally, two sets of the stance phase comprised eight legs alternately supporting and driving the motion of the spider’s body. The spider primarily increased its movement velocity by increasing stride frequency. In comparison to other insects, the spider, O. [sic] huwena, has superior movement stability. The velocity and height of COM periodically fluctuated during movement, reaching a maximum during alternation of leg phase, and falling to a minimum in the steady stance phase. The small change in deflection angle of the hind-leg was effective in driving locomotion, whereas each joint-rotation angle of the fore-leg changed irregularly during locomotion. This research will help in the design of bio-inspired robots, including the selection of gait planning and its control.
Machkour-M’Rabet, S., Y. Hénaut, S. Calmé & L. Legal. 2011. When landscape modification is advantageous for protected species. The case of a synanthropic tarantula, Brachypelma vagans. Journal of Insect Conservation. Online publication date: 24-Aug-2011. DOI: 10.1007/s10841-011-9434-3
Abstract. Landscape fragmentation usually has a considerable effect on the genetic and demographic viability of most species because it reduces population size and increases isolation among populations. This situation provokes loss of genetic diversity and increased inbreeding that can lead to population or species extinctions. Some studies also show that landscape fragmentation may have no effect on or even positive consequences for species genetic diversity. The protected tarantula, Brachypelma vagans, exhibits a particular situation in the Mexican Caribbean, which has experienced high lowland and coastal fragmentation because of recent increases in agricultural, urban and touristic development. This modified landscape structure creates favorable conditions for establishment of B. vagans populations in rural settlements. Populations of this tarantula have high densities of individuals, principally females and juveniles, and gene dispersion is assumed by the rare males. Within this context, we studied the influence of natural and anthropogenic fragmentation on the genetic diversity of six B. vagans populations (five continental, one insular), together with their spatial organization. Our approach used seven inter simple sequence repeat markers, which are highly polymorphic markers. The 76 loci selected revealed high genetic variability for continental populations and a low, but not critical situation, for the insular population. We detected a good level of gene exchange among continental populations, and an evident and recent isolation of the island population. This species exhibits a metapopulation structure in the lowlands with numerous local populations where mature females exhibit high birth site fidelity. We conclude that this protected species does not exhibit characteristics to warrant its current conservation status, and we propose complete revision of the ecological and genetic situation for B. vagans in particular, and for all species within the genus Brachypelma in general.
Tuesday, August 02, 2011
12. Pokasztalalkozo
Kedves Pókászok és Pókbarátok!
Köszönjük mindazoknak, akik válaszoltak és támogatták javaslatunkat, miszerint 2011-ben Zala megye adjon otthont a rendes évi Magyar Pókász Találkozónak.
Örömmel tudatjuk Veletek, hogy a mára már hagyományossá vált, hazai pókász találkozónak tehát a dimbes-dombos, ezer éves táj: Zala megye ad otthont.
Mindenkit szeretettel várunk a Zala – völgyébe, Pakodra, ahol lehetőségünk nyílik szakmázni, megismerkedni a környékkel és remélhetőleg jó hangulatban, együtt eltölteni pár napot.
A találkozó tervezett időpontja: 2011. szeptember 16-18, helyszíne Zala megye, Pakod: Endes major.
A szállásról bővebben tájékozódni, fényképeket nézegetni a http://www.endesmajor.fw.hu oldalon tudtok.
A tervezett programokról, várható költségekről, illetve a jelentkezési és egyéb határidőkről az augusztus eleji hírlevelünkből értesültök majd.
Addig is kívánunk mindenkinek további szép nyarat, kinek pihenést, kinek szerencsés terepnapokat!
Zalaegerszeg, 2011. július 28.
Kovács Péter és Kancsal Béla
a szervezők
Köszönjük mindazoknak, akik válaszoltak és támogatták javaslatunkat, miszerint 2011-ben Zala megye adjon otthont a rendes évi Magyar Pókász Találkozónak.
Örömmel tudatjuk Veletek, hogy a mára már hagyományossá vált, hazai pókász találkozónak tehát a dimbes-dombos, ezer éves táj: Zala megye ad otthont.
Mindenkit szeretettel várunk a Zala – völgyébe, Pakodra, ahol lehetőségünk nyílik szakmázni, megismerkedni a környékkel és remélhetőleg jó hangulatban, együtt eltölteni pár napot.
A találkozó tervezett időpontja: 2011. szeptember 16-18, helyszíne Zala megye, Pakod: Endes major.
A szállásról bővebben tájékozódni, fényképeket nézegetni a http://www.endesmajor.fw.hu oldalon tudtok.
A tervezett programokról, várható költségekről, illetve a jelentkezési és egyéb határidőkről az augusztus eleji hírlevelünkből értesültök majd.
Addig is kívánunk mindenkinek további szép nyarat, kinek pihenést, kinek szerencsés terepnapokat!
Zalaegerszeg, 2011. július 28.
Kovács Péter és Kancsal Béla
a szervezők
Monday, July 04, 2011
Preparálás I. A szárazpreparátum

Terveim szerint ebben a sorozatban időről - időre ismertetve lesz egy-egy módszer, ahogyan preparátumot készíthetünk az elpusztult madárpókokból vagy azok egyes részeiből. E sorozat első része a szárazpreparátum készítése, amely talán a „közember" által leggyakrabban alkalmazott preparálási technika ha egy ízeltlábúról van szó.
Csak vázlatpontszerűen értekeznék az általam alkalmazott eljárásról, mellyel szárazpreparátumot készítek az elpusztult nagyobb termetű madárpókokból, mivel nem hinném, hogy sok új információt adok közre ez ügyben. Ellenben lehet, hogy valakinek éppen az alább leírtak hiányoznak a boldog hétköznapokhoz (who knows?)…vágjunk is bele:
A művelet egyszerű, mint a barlangrajz, nem kell semmiféle firlefáncos űrtechnológiára gondolni. Persze ízlés szerint lehet cizellálni mindenféle kegyszerrel és vegyszerrel, meg festett műutótesttel..stb, így aki nem elégszik meg ezzel az egyszerű mezei eljárással, ami azért a bazárokban kapható „The Spider” cimkével ellátott förmedvényeket még mindig fényévekkel előzi, teljes nyugodtsággal tanulmányozza
Dr. Fehér György könyvét a tökéletes nirvána elérésének érdekében.
A demonstrációs célra felhasznált állat egy Pamphobeteus nigricolor kifejlett nőstény egyede, mely már jó ideje mélyhűtött állapotban „várakozik” postmortem állapotban a frigóban. Érdekesség kedvéért, így nézett ki ereje teljében (amint épp "kicsontozta" a vőlegényét):

Az alábbi képhez jelen esetben nem fűznék a korábbiakban alkalmazott számozott szájbarágót. Szerintem elég egyértelműen beazonosíthatók a jószág kikészítéséhez használt eszközök. Ha mégsem, akkor azt is ki tudjuk tárgyalni komment jelleggel. Sajnos a képről lemaradt, de mindenképpen említést érdemel egy formalinnal töltött injekciós fecskendő. Erre később még kitérek.
Éles penge segítségével, hosszanti irányú vágással nyissuk fel az utótest hasi oldalát.
Majd távolítsuk el az utótest tartalmát. Ha ezt nem tesszük, akkor egyrészt a száradás végeredményeképpen az utótest összezsugorodik, másrészt az utótest tartalmának rothadása következtében terjengő bűz miatt a pók nem igazán lesz alkalmas lakásdekorációs célokra (szóltam!:) ).
E célra nekem legjobban a hurkapálcára fordított csepp alakban felcsavart vatta vált be a legjobban, melyet csavaró mozdulatokkal alkalmazok az utótest tartalmának eltávolítására. Ezután addig kell cserélni a vattalabdacsokat, amíg szinte szárazra töröljük az utótest belsejét, és semmiféle elszíneződés nem látszik a felvágott utótestből kihúzott vattán. Legvégül alkohollal vagy formalin oldattal mossuk végig az utótest kültakarójának belső oldalát, majd ismét töröljük szárazra.

A kitöméshez szintén vattát használok. Nagyjából borsó nagyságú golyókat formázok vattából és fokozatosan töltöm fel velük az utótest kültakaróját. Tapasztalataim szerint ezáltal sokkal könnyebb feszesebbre tömni, és végül megfelelő formára alakítani az utótestet, mintha nagyobb darabokkal járnánk el ugyanígy. A kitöméshez használt vattát érdemes alkoholba vagy formalin oldatba mártani a további baktériumos folyamatok csökkentésének érdekében. Én nem használok, de biztos nem árt ha ennél a műveletnél is alkalmazunk rovarmérgeket a kártevők ellen (múzeumbogár, moly..stb.).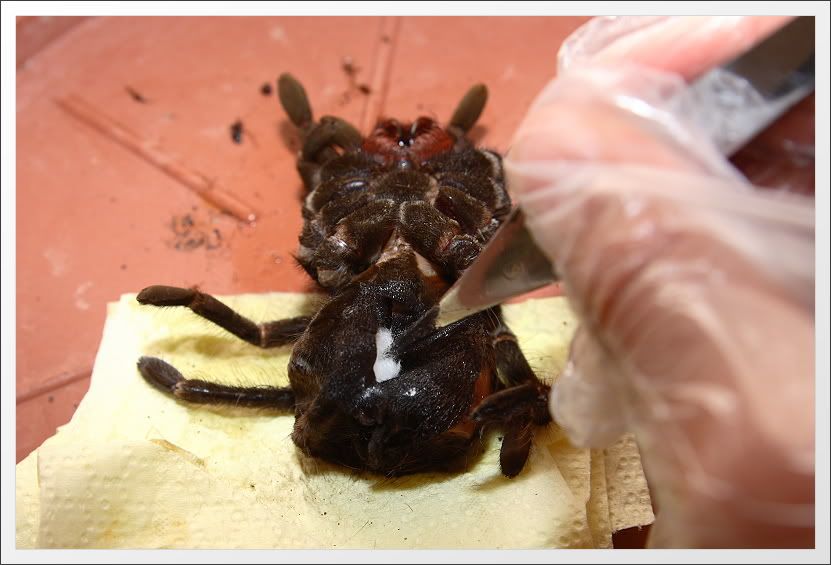
A művelet végén finoman a kívánt alakúra lehet gyurmázni (ujjbeggyel nyomkodni) az utótestet.

Ahogy a képen is látszik, a fagyasztással történő tárolás közben megsérülhet a madárpók. Főképpen a lábak törnek könnyen. A szükséges ragasztást ennél a műveletnél végzem, a következő fotón látható technika alkalmazásával.
Egyszerű „toldót” faragok gyufaszálból, fogpiszkálóból..stb, hogy a ragasztás stabilabb legyen.
Ezután következik az állat rögzítése a kívánt testtartásban. Jelen esetben a pókot általános pihenő pozitúrában rögzítettem, enyhén terpesztett lábakkal. Ehhez tökéletesen megfelel a háztartásokban található, olcsó gombostű.

A fonószemölcsök rögzítése:
Amikor a rögzítéssel kész vagyok, vékony injekciós tű segítségével, a képen jelölt helyen, az izületi membránon keresztül formalin oldatot fecskendezek a lábakba, a baktériumos dzsembori csökkentésének céljából. Főképpen a lefagyasztott, majd felengedett példányok esetében javallott megtenni, mivel a fagyasztás következtében „megfolyósodott” lágy szövetek „beindulva” irgalmatlan bűzt képesek produkálni. A formalin használatakor mindenképpen érdemes szem előtt tartani, hogy egy olyan méreg, ami bőrre kerülve vagy belélegezve elég kellemetlen, de ráadásul legalább rákkeltő is (munkavédelmi előírások betartása éppen ezért javallott:) ).
Amikor a preparált példányunk tökéletesen megszáradt egy fóliával letakarva megóvhatjuk a porosodástól, amíg döntünk a továbbiakról.
A preparált madárpók végleges tárolására és bemutatására legcélszerűbb egy rovardobozt beszerezni. Ennek hiányában megfelel bármilyen magasabb oldalfallal ellátott jól záródó doboz, amely megóvja a preparátumot a nemkívánatos külső behatásoktól (kártevők, por…stb).
Erre a célra ezt az Ikeában kapható dobozt találtam a legmegfelelőbbnek, amelyet sajnos máshol még nem láttam (hasonlót sem), így a beszerzése vidékről legalább olyan macerás, mint egy rovardoboznak.
Legvégül következik a szárazpreparátum montírozása. A pók végízeinek talpecsetét pillanatragasztóval a talapzathoz rögzítem, majd megkapja az elmaradhatatlan címkét.
Kész, mehet a dobozba:
Élőben sokkal „nagyobbat” mutat:)
Ha van kérdés, nyugodtan jöhet itt komment formában, de a Facebookon is!
Tuesday, June 28, 2011
Díszítősor: Durcás szépségek

A napokban aktuális, hosszabb lélegzetvételű bejegyzésem előtt, néhány fotó ezekről a durcás szépségekről. Zolinak ajánlom.
0.1. Xenesthis sp. „blue”
Spermatartó:
0.1. Xenesthis immanis

Spermatartó:
Sunday, June 19, 2011


Friday, June 17, 2011
Majka a dajka

Arra azért kiváncsi lennék, hogy miként került a lakásba egy Cheiracanthium… Azért furcsálom a dolgot, mert itt a környéken (Szekszárd) nem túl gyakori ez a pók. Legalábbis idáig még nem találkoztam vele errefelé.
A parkettán sietős léptekkel tovatűnő, viszonylag nagy méretű, nyurga jószágról először azt gondoltam, hogy egy „betévedt” bolygó hollandi Tegenaria hím. Mivel az életteremben nem kedvelem a spontán szerveződő biodiverzitást (vannak itt elegen tervszerűen:) ), ezért gyorsan ráborítottam a kezem ügyében levő Sunpak diffúzort, nehogy a kis delikvensünk a váratlan helyzetet kihasználva olajra lépjen. Legnagyobb meglepetésemre az alkalmi kupola alatt sacperkábé hiperűrsebességgel elkezdett pattogni, mint egy megbokrosodott nikkelbolha. A rögtönzött rodeo akkora beleéléssel zajlott, hogy a nagy mutatvány közben, a jobb I. járóláb is kótyavetyére került, ezáltal jól tanulmányozhatóvá vált az autotomiás pont.
Az előállítást követő rabosításnál viszont én képedtem el, látva, hogy ki rajcsúrozott itt a lábam mellett.
Most azon tűnődöm, hogy mi legyen a sorsa ennek a kis negatív hősnek, akit sebtében Majkának neveztem el (ez jutott eszembe erről az arcszerkezetről)…
Wednesday, June 15, 2011
Bazár bezár

Vagyis a DNFS fórum hamarosan végleg lekapcsolódik. Alábbiakban olvasható ZoozDa közleménye:
"Sziasztok,
fájó szívvel , de 2011 jún. 15.-én bezár (eltűnik) az oldal. Sajnos a domaint kezelő cég csak most szólt, így nincs sok idő variálni.
Részemről nem finanszírozom tovább a domaint évi 10k forintért ért (csak a tárhely volt ingyen) úgy, hogy 4-5 ember hírdetésén
túl nem sok minden történt itt az utóbbi 2-3 évben.
Ráadásul én magam is megválok/váltam a pókjaimtól, és nem is tervezem a közeljövőben a velük való foglakozást,
még sokadik hobbyként sem.
Mindenkinek további jó pókozást kívánok.
Üdv,
ZoozDa"
Wednesday, June 08, 2011
5 random régebbi madárpókos cikk
Pocock, R. I. 1903. On some genera and species of South American Aviculariidae. Annals and Magazine of Natural History (7) 11: 81–115. Kattints ide a letöltéshez!
Koch, C. L. 1841–1842. Die Arachniden. Nürnberg, Neunter Band, 108 pp. Kattints ide a letöltéshez!
Pocock, R. I. 1902. Some new African spiders. Annals and Magazine of Natural History (7) 10: 315–530. Kattints ide a letöltéshez!
Mello-Leitão, C. F. de. 1923. Theraphosideas do Brasil. Revista do Museu Paulista 13: 1–438. Kattints ide a letöltéshez!
Chamberlin, R. V. 1917. New spiders of the family Aviculariidae. Bulletin of the Museum of Comparative Zoology at Harvard College 61: 25–75. Kattints ide a letöltéshez!
Monday, June 06, 2011
A pókok fő csoportjairól I.
A pókok (Araneae) napjainkban elfogadott magasabb (alrend és osztag) szintű osztályozását először Pocock javasolta 1892-ben.


Az Mesothelae alrendet egyetlen család (Liphistiidae – félpókok) 5 génuszának 89 faja alkotja (Platnick, 2011). Ők a többi pók – Opisthothelae = Mygalomorphae és Araneomorphae osztagok együtt – testvércsoportja. A Liphistiidae, Mygalomorphae és Araneomorphae rokonsági viszonya olyan, mint az unokatestvéred (L), a testvéred (M) és te (A). Az Opisthothelae az utolsó WSC verzió szerint 109 családot, 3816 génuszt és 41966 fajt jelent, de azóta már sok tudományra új pókfajt leírtak (csak madárpókfajt négyet). Persze ezt úgy kell értelmezni, hogy ezek a jelenleg érvényes nominális fajok és génuszok. Új taxonok leírása nélkül, a jelenlegiek átvizsgálásával ez a szám is változna, mivel jó eséllyel új szinonimákat állítanának fel, vagy régieket nem fogadnának el stb.
A XX. században Pocockén kívül több különböző klasszifikációt publikáltak különböző arachnológusok (pl. Petrunkevitch, Bristowe, Millot, Gertsch). Talán Platnick & Gertsch (1976) közölték az első olyan tanulmányt, ami az első kladisztika volt pókokon, vagyis azzal a kritériummal dolgoztak, hogy minden megnevezett taxon monofiletikus kell, hogy legyen. Eredményeik megegyeztek Pocockéval (1892). Egy taxont monofiletikusnak mondunk, ha egy feltételezett közös ős összes leszármazottját magában foglalja. A kladisztikában egy monofiletikus csoportot a csoport tagjaira jellemző közös levezetett jellegek (szünapomorfiák) alapján állítják fel, szemben az ősi (primitív) közös jellegekkel (szünpleziomorfiák). Platnick & Gertsch (1976) négy (4) karaktert javasoltak, amik a Mesothelae alrend feltételezett apomorfiái, és ezáltal a közös őstől való leszármazásukat támasztják alá. Megjegyzés: a többször előforduló összehasonlítás az ostorlábúakkal azért van, mert őket tartják a pókok legközelebbi rokonának, így Platnickék őket használták külcsoportként.
(1) A negyedik lábak csípőjén található betűrődés (invagináció). A többi póknál és az ostorlábúaknál (Amblypygi) ez nincs meg.
(2) A Heptahela kimurai haploid kromoszóma száma 48, míg a Platnickék által vizsgált pókoké 24 vagy kevesebb, az ostorlábú Sarax sarawakensis-nél pedig szintén 24 haploid kromoszómát mutattak ki.
(3) A trichobothriumok (érzékelő képletek) töveinek szerkezete a lábak disztális ízein különbözik a liphistiidáknál a többi pókkal összehasonlítva. A liphistiidáknál a trichobothrium egy kupolaszerű szerkezetből lép ki a testből, két oldalról egy-egy lemezzel határolva. A Platnick & Gertsch (1976: 10, 9. és 11. ábra) által közölt képeknél a 9. ábrán (Liphistius sp.) ez tisztán látszik, azonban a 11. ábrán (Heptathela sp.), a kupola mélyen helyezkedik el és én személy szerint csak az egyik lemezt látom. Ez lehet a scan szöge miatt is, azaz amiatt, hogy nem egy síkban mutatja a lemezeket. Haupt (2003) állítása szerint csak egy lemezt talált az általa vizsgált liphistiid fajoknál, bár ő ugye méregmirigyeket sem talált, Foelix & Erb pedig igen. N. I. Platnick szerint (személyes közlés) a liphistiidák trichobothrium töveinek felépítését nem lehet összetéveszteni más pókokéval („I don't recall seeing any trichobothrial bases in other spiders that could be confused with the liphistiid arrangements”). A többi póknál egy lemez vagy egyszerű kupola figyelhető meg. A szerzők mindkét állapotot levezetettnek tekintik.
(4) Az oldalsó (laterális) fonószemölcsök felépítése különböző a pókok három fő csoportjánál. A Liphistiidae családnál sok szegmensből állnak (multisegmented), akár 12-ből is. A Mygalomorphae poszterior laterális fonószemölcsei három vagy négy szegmensből állnak, míg az Araneomorphae-nél egy vagy két szegmens alkotja őket. Ezen karakternél nem tudták megállapítani a polaritást, azaz, hogy melyik állapot az ősi és melyik a levezetett. A sokszegmensű állapot tehát a Mesothelae, a négy vagy kevesebb szegmensű állapot pedig az Opisthothelae monofiletikusságát támaszthatja alá.
Később Platnick & Goloboff (1985) egy azelőtt ismeretlen jellegről számoltak be, ami kizárólag a Mesothelae-ra jellemző, így szintén az alrend közös őstől való leszármazását igazolja:
(5) Az I-III. lábak lábszárának pro- és retrolaterális oldalán egy-egy sarkantyút találtak, amik az I-III. lábak másodvégízének tövi részénél levő, enyhén emelt, nem megkeményedett területtel lehetnek képesek érintkezni. A karaktert ivaréretlen és ivarérett nőstények, valamint penultimate hímeknél figyelték meg, az ivarérett hímek viszont úgy tűnik, elveszítik ezt.
Raven (1985) egy további apomorfiát javasolt:
(6) A Mesothelae-nél a mellpajzs (sternum) vékony és kiemelkedik a testből, egy meredek szélű fennsíkra emlékeztet. Az Opisthothelae-nél viszont a sternum sokkal szélesebb és laposabb. Ellenben a legtöbb pókszabásúnál, így az ostorlábúaknál is (pókok testvércsoportja) a mellpajzs hiányzik vagy gyengén fejlett. Raven (1985) szerint a pókoknál megfigyelhető mindkét állapot az adott csoport levezetett jellegének tekinthető.
A fent és lentebb tárgyalt Liphistiidae tulajdonságok egy része megfigyelhető ezeken a képeken: kép 1, kép 2. A félpókokra jellemző feltűnő tulajdonság az utótesten a szegmentáltság nyomát jelképező lemezek (tergite),, illetve a fonószemölcsök száma, még inkább azok pozíciója, amiket a többi pókkal ellentétben az utótest hasi oldalának közepe fele, és nem terminális (vég) pozícióban találunk meg. Mivel mindkettő ősi jelleg/állapot, azonosítás szempontjából hasznosak lehetnek, de nem használhatóak a csoport monofiletikusságának megállapítására.
A következőkben tekintsük át az Opisthothelae (Mygalomorphae + Araneomorphae) levezetett karaktereit.
(1) (2) Ld. Mesothelae-nél a 3. és 4. jellegek.
(3) A liphistiidáknak 8 vagy 7 fonószemölcsük van, a többi póknak 6 vagy annál kevesebb. Ismert, hogy a szövőlap és a colulus az aranemorfoknál az egyedfejlődés során a két „hiányzó” elülső középső fonószemölcs egyesüléséből jön létre, ezen fonószemölcsök elvesztését levezetettnek tekintik.
(4) Ahogy kicsivel feljebb szerepel, a liphistiidáknál a fonószemölcsök az utótest hasi oldalának közepén vannak. A többi póknál a kaudális (farok) szegmensek az egyedfejlődés során egyesülnek, és a fonószemölcsök az utótest végéhez (és az ánuszhoz) kerülnek közel, ezért utóbbi állapot az apomorf.
(5) A liphistiidáknál egy kis, kerek sclerit lemez formájában megmarad az utótest első szegmensének hasi lemeze (sternite). Mivel az ostorlábúak is megőrzik ezt egy többé-kevésbé megkeményedett képlet formájában a pedicelen, a többi póknál az első sternite teljes redukcióját levezetettnek tekintik.
(6) A félpókok második pár tracheatüdőjét viselő harmadik szegmense az utótestnek hasi oldalról egy határozott sclerit lemez. Ez a sternite az ostorlábúaknál is világosan jelen van, a szegmentáltság eme külső jelének elvesztése a többi póknál egy újabb apomorfia.
(7) A subesophageális (nyelőcső alatti) ganglionok a Millot által vizsgált ostorlábúaknál és négy Liphistius fajnál 17 neuromert tartalmaznak, míg a többi póknál 12-t, a kisebb számot vélik levezetettnek.
(8) Ld. Mesothelae-nél 6. jelleg.
Egy közeljövőbeni bejegyzésben hasonló stílusban a Mygalomorphae és Araneomorphae közötti különbségeket, illetve levezetett jellegeiket fogjuk ismertetni.
Irodalom
1. Haupt, J. 2003. The Mesothelae -- a monograph of an exceptional group of spiders (Aaneae: Mesothelae): (Morphology, behaviour, ecology, taxonomy, distribution and phylogeny). Zoologica 154: 1–102.
2. Pocock, R. I. 1892. Liphistius and its bearing upon the classification of spiders. Annals and Magazine of Natural History (6) 10: 306–314.
3. Platnick, N. I. 2011. The world spider catalog, version 11.5. American Museum of Natural History, online at
http://research.amnh.org/iz/spiders/catalog. DOI: 10.5531/db.iz.0001.
4. Platnick, N. I. & W. J. Gertsch. 1976. The suborders of spiders: a cladistic analysis (Arachnida, Araneae). American Museum Novitates 2607: 1–15. URI: http://hdl.handle.net/2246/5468
5. Platnick, N. I. & P. A. Goloboff. 1985. On the monophyly of the spider suborder Mesothelae (Arachnida: Araneae). Journal of the New York Entomological Society 93 (4): 1265–1270.
6. Raven, R. J. 1985. The spider infraorder Mygalomorphae (Araneae): cladistics and systematics. Bulletin of the American Museum of Natural History 182: 1–180. URI: http://hdl.handle.net/2246/955
Sunday, May 29, 2011
A vészfék

A többi pókhoz hasonlóan a madárpókok (Theraphosidae) is állítanak elő pókselymet az erre szolgáló speciális mirigyeikben, amit az utótestükön található fonószemölcsökből juttatnak a külvilágba. Korábban felmerült, hogy az egyik nagytermetű madárpókfaj, az Aphonopelma seemanni képes pókselymet kiválasztani a járólábainak végízeiből, bár ez az állítást Perez-Miles és munkatársai később megkérdőjelezték, mivel az erre irányuló kutatás nem vezetett eredményre.

Brachypelma auratum függőleges falat mászik
Ebben a cikkben (Rind et al. 2011. Tarantulas cling to smooth vertical surfaces by secreting silk from their feet. The Journal of Experimental Biology 214, 1874-1879.) a szerzők bizonyítják, hogy olyan szövőcsévék találhatóak három közismert madárpókfaj (Grammostola rosea, Poecilotheria regalis, Brachypelma auratum) járólábainak végízén, amelyek közvetlenül képesek pókselyemet létrehozni.
Brachypelma auratum függőleges falat mászik, alulnézet
Az előzőekben említett fajok alapján arra lehet következtetni, hogy ez a képesség széles körben elterjedt a madárpókoknál. A cikk bemutatja, ahogyan a madárpók egy több szálból álló „hálólábnyomot” hoz létre, amikor elkezd lefelé csúszni egy sima, vízszintes felületen. A végízen található kellően erős és rugalmas fúvóka szerű szőrök felelősek a pókfonal „ledobásáért”, melyek végül „lehorgonyozzák” a madárpókot.
Grammostola rosea selymet választ ki a lábából. A) A példány a függőleges falon mászik; B) Selyem a falon; C) A selyem a felületen, 27 szál; levedlett tapogatóláb rétegfotója.
A szerzők utalnak arra is, hogy a madárpókok fonószemölcsein található szövőcsévék nagyon hasonlítanak a Devon korból ismert ős-pók fonószemölcsére. Ez arra enged következtetni, hogy a pókselyem kiválasztásának mikéntje a madárpókoknál igen ősi formában maradt fent.
A-B) Grammostola rosea lábvégíz, talpkefe, szkenning elektronmikroszkópos kép.
C) Grammostola rosea részlegesen levedlett láb keresztmetszete; D) Brachypelma auratum lábvégvedlés belülről a szövőcsévéket nyilak jelzik.
Poecilotheria regalis lábvégi szövőcsévék, szkenning elektronmikroszkópos képek.
Forrás:
Rind, C., Birkett, C. L., Duncan, B.-J. A. and Ranken, A. J. Tarantulas cling to smooth vertical surfaces by secreting silk from their feet.. The Journal of Experimental Biology, 214, 1874-1879 DOI: 10.1242/jeb.055657

{kind=link}
Wednesday, May 25, 2011
Nevén a gyereket

A Jurai arany selyempók bejegyzésben Tamás azt írta, hogy a MYGALOMORPHAE Pocock, 1892: 311 infraordonak „még mindig nincs kielégítően jó magyar neve.” Ha megnézzük Loksa Imre Pókok I. magyar nyelvű határozókönyvét, akkor azt látjuk, hogy a „négytüdős” kifejezést használta a magyar aknászpókra és a három torzpókfajra, akik a Mygalomorphae magyar képviselői. Elképzelhető, hogy ez egyszerűen a régebbi név TETRAPNEUMONES fordítása (a korai rendszerezésekben a másik csoport neve DIPNEUMONES – kéttüdősök volt). A tudományos neveket félretéve a „négytüdős pókok” nem logikus, mert
a.) van olyan mygalomorph taxon, aminek nincs egyetlen tracheatüdője sem, a Micromygale diblemma Platnick & Forster, 1982 (a Micromygale monotipikus génusz);
b.) van olyan nem-mygalomorph taxon, aminek négy tracheatüdője van, pl. az Austrochiloidea és Hypochiloidea családsorozatok (nem Onedin vagy Ewing féle családsorozat): Hickmania (Austrochilidae, a másik két austrochilid génusznál a hátsó pár tracheatüdők módosultak), Hypochilidae, Gradungulidae családok (Araneomorphae); Liphistiidae család, utóbbi képviselői nem mygalomorfok, hanem a legősibb pókokként az összes többi pók testvércsoportját alkotják a Mesothelae alrendként (a testvércsoport Opisthothelae = Mygalomorphae + Araneomorphae). A Hypochilidae stb. családok tüdőinek száma ellenére közelebbi rokonságban áll a „kéttüdősökkel” (araneomorfok), mint a mygalmorfokkal.
A pókok esetében többször végbement a tracheatüdők redukciója pl. a Haplogyne csoport esetében (szintén nincs kielégítő és jó magyar nevük) ahol max négy trachea található, illetve a főpókoknál a két tüdő egy tracheanyílásból elágazó légcsőrendszer az általános.
A pókok magasabb szintű osztályozásába nem akartunk itt belemenni (akit érdekel, ezt töltse le), így maradjunk annyiban, hogy a „négytüdős” név logikailag azért nem jó, mert nem minden négy tracheatüdővel rendelkező csoport tartozik ide, és a csoportnak nem minden tagja rendelkezik négy tracheatüdővel. Persze tudjuk, hogy a nyelvben olykor nem a logika, hanem a hagyomány az irányadó, így kapóra jön, hogy a Mygalomorphae is tulajdonképpen egy hagyományőrző név. A fentebb leírtak miatt látta Pocock szükségesnek a Tetrapneumones és Dipneumones nevek elvetését, és helyettük a Mygalomorphae és Arachnomorphae kifejezéseket javasolta. Utóbbi tartalmában megegyezik, írásában eltér a ma használt Araneomorphae-val.



A Mygalomorphae név választása mögött pedig a következő áll. Eleinte az összes pókot az Araneus ill. Aranea generikus név alatt tartották számon. Latreille (1802, 1804) Mygale génusza volt az első olyan génusz (nevezzük itt most csoportnak), ami a pókokat kettéosztotta, illetve elkezdte megkülönböztetni egymástól. Latreille 1804-es publikációjában a következő két fajt írta le és sorolta ide: Mygale blondii és Mygale fasciata (mai nevükön Theraphosa blondi és Poecilotheria fasciata). Ezt követően több későbbi mygalomorph pókot a Mygale génuszban írták le. Sajnos a Mygale név pókokra használva nem maradhatott fent (Thorell, 1870), mivel még Latreille előtt használta egy másik szerző (Cuvier, 1800) egy emlősgénusz neveként, ld. dezmán – Mygale moschata (a mygale görög jelentése is elvileg „shrew” = cickány). Ezzel együtt a Mygale nevet a köztudat ezekkel a „nagy és szőrös” pókokkal azonosította, és sokáig így ismerték őket a „be nem avatottak”, valamint sok zoológiai és természetrajzi könyvbe is ilyen néven kerültek be. Pocock ezért válaszotta a Mygalomorphae nevet („Mygale-alkatúak”), és mivel a csoport talán legismertebb és legjellegzetesebb képviselői a ma madárpókokként ismert Theraphosidae család tagjai (mellesleg a legelső Mygale-ként leírt fajok is madárpókok voltak), ezért szerintünk a „madárpókalkatúak” vagy „madárpókfélék” jó választás lenne a Mygalomorphae magyar neveként.
Irodalom
1. Latreille, P. A. 1802. Histoire naturelle, générale et particulière des Crustacés et des Insectes. Paris, 7: 48-59.
2. Latreille, P. A. 1804. Histoire naturelle, générale et particulière des Crustacés et des Insectes. Paris, 7: 144-305.
3. Platnick, N. I. & R. R. Forster. 1982. On the Micromygalinae, a new subfamily of mygalomorph spiders (Araneae, Microstigmatidae). American Museum Novitates 2734: 1–13.
4. Pocock, R. I. 1892. Liphistius and its bearing upon the classification of spiders. Annals and Magazine of Natural History (6) 10: 306–314.
5. Thorell, T. 1870. On European spiders. Nova Acta Regiæ Societatis Scientiarum Upsaliensis (3) 7: 109–242.
Üdv: Zoltán & siler.
Tuesday, May 17, 2011
Conica Minolta

Vannak lehetetlennek tűnő műveletek, melyektől hamar ledobja az agyam a szíjat. A Csúcsos keresztespók (Cyclosa conica) és más hasonszőrű, liliputi méretű kerekhálósok fotózása terepen számomra többnyire ebbe a kategóriába tartozik.
Mivel a korábbi próbálkozásaim eredményétől nem vágtam magam hanyatt, úgy döntöttem, hogy ennek az érdekes apró póknak a fotózását illetően futok még egy kört.
Az advanced szintű kihívás az állat életmódjában leledzik, melyet Dr. Szinetár Csaba fogalmaz meg roppant frappánsan a Pókok című könyvének 52. oldalán: „Kedveli az elszáradt ágakból álló, kevéssé zárt légifolyosókat”.
Na igen! Ezekben a kevéssé zárt légifolyosókban, napközben a szélcsend a legritkább jelenség, így mialatt a gép keresőjén keresztül megpróbálunk ráfókuszálni a háló közepén héderező pókra, hasonló látványban van részünk, mint amikor a halálcsillag megtámadásánál Kardfater próbálta befogni a TIE vadász célkeresztjébe az ereje teljében levő Skywalker X-szárnyúját. Vagyis…ezt a könnyű kis jószágot a leglágyabb nyári szellő is random képes belengetni az összes axonometrikus tengely mentén, pont olyan formán, mint amit Keith Flinttől láthattunk a Firestarter c. nótázós Prodigy klippben.
Így néz ki egy „kevéssé zárt légifolyosóban” elkészített kerekháló, tokkal vonóval, ami esetünkben egy fa lelógó ágvégei között lett kifeszítve, és a pókon kívül még az állat közelmúltban elhíresült „árnyékra ugrató álcázóberendezését ” is magába foglalja.
Ennek ellenére némi türelemmel, szerencsével és erős mesterlövész képességekkel felvértezve azért létre lehet hozni néhány elfogadható minőségű fotót, de 1:1 közelében így sem nagyon kell egerészni.

Jó, elismerem, hogy tűhegynyi blendenyílással meg egy atomvillanással még az előttünk levő 1 hektáros búzatábla legtávolabbi vége is éles lesz, de én szerettem volna olyan minőségi képet erről a pókról, amin a háttér is kap fényt, nemcsak olyat ahol a kizárólag a téma. Mindemellett megfelelő mélységélesség tartománnyal garázdálkodhatok bemozdulásmentesen.
Ilyenkor kérdezi a népi folklór, hogy „namég a kis @-om nem kéne”? De No Problémo, mert ehhez nem kellett mást tennem, mint a pókkal egy fotózásra alkalmas, szélcsendes helyen megszövetni a hálóját. Fogtam tehát és hazavittem. Eszkábáltam neki egy ideális póklakot, melyen kedvemre megfigyelhettem és fotózhattam. Az első két napot átsziesztázta, csak éjszaka tevékenykedett, majd a harmadik napon megszőtte a hálóját.

Ezután következhetett a fotózás, „háttérfénnyel”,

…és háttérfény nélkül.

Egy kereresztespókok fotózására alkalmas megfigyelő setupot könnyűszerrel össze lehet dobni. Hardcore hobbisták akár a pókos szobájukat is dekorálhatják vele, de az biztos, hogy a pókoktól rettegő családtagokban és betérő ismerősökben nem arat osztatlan sikert pl. egy monitor mellett felállított dizájnos „szabadtéri” predátor kuckó.
Én ezekből az elemekből legóztam össze:
1, Virágcserép (lehetőleg kerámia, mert az nehezebben borul)
2, Tálca
3, Villás faág
4, Papír
5, Gyurma
A cserépen keresztülbújtatott ágat egy kicsit lesúlyozom gyurmával, hogy még stabilabban álljon.
A cserép nyílásának vastagságához passzinthatjuk az ágat némi rátekert papír segítségével.
Ezután feltöltjük a tálcát vízzel. Fontos, hogy a tálca szélesebb legyen, mint az ágvilla, tehát a leereszkedő pók mindig csak a vízbe tudjon érkezni, különben néhány Houdini tulajdonsággal megáldott faj nemes egyszerűséggel meglép (a Cyclosa conica nem annyira szökős).
Legvégül jöhet valamelyik rosszarcú… jelen esetben egy Kétcsúcsú keresztespók (Araneus bituberculatus), aki a befogását követően fellépő heveny fiolahiány miatt kénytelen volt együtt utazni néhány Lampropelma-nak szánt szövőlepkével. Ez a magyarázata annak, hogy némi hímpor látható az ábrázatán.

Mivel nem vagyunk kitéve semmiféle zavaró időjárási tényezőnek, bekvártélyozás után már melegében „meg is lőhetjük” néhány próbafotó erejéig az újdonsült delikvensünket:

Subscribe to:
Posts (Atom)